

















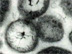










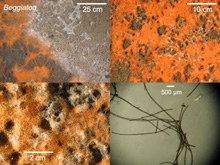
Fig. 1. Beggiatoa mats on the sea floor around a Gulf of Mexico cold seep. The image mosaic (from left to right, and top to bottom) shows increasing magnification of the individual filaments, which are clearly visible in the 2-cm scale bar panel. Click image for larger view and image credit.
Microbiology
Mandy Joye
Professor of Marine Sciences
University of Georgia
Introduction
Microbial populations, representing both Bacteria and Archaea, are extremely diverse. Though most individual microorganisms are invisible to the naked eye, in total they account for about half of the carbon in organic biomass on the Earth (Whitman et al. 1998). Yet, their diversity and functional role in many habitats are still poorly understood. This is particularly true for deep-ocean cold seeps.
Life Without Sunlight
How is it possible for life to flourish on the dark, deep sea floor? At the surface of the Earth, sunlight fuels photosynthesis, a process by which primary producers, like tomato plants in your garden or microscopic algae in lakes, use energy captured from the sunlight to transform carbon dioxide into organic carbon (i.e., plant biomass). This plant biomass is consumed by other organisms and the energy is transferred up the food web to higher organisms. “Primary” production forms the base of the food chain.
In the deep ocean, there is no sunlight (and therefore no photosynthesis), yet life flourishes in certain places. Sea-floor cold seeps are just such places. Cold seeps are characterized by large amounts of energy-rich compounds (e.g., hydrogen sulfide, methane, and ammonium) that travel from deep reservoirs, seeping up to the sea floor. These compounds are used by microorganisms called chemoautotrophs in a process called chemosynthesis. Chemoautotrophs use energy obtained from the oxidation of hydrogen sulfide, methane, or ammonium, to transform carbon dioxide into organic biomass. These microorganisms play an ecological role similar to photosynthetic plants, in that they serve as the base of the food chain. At deep-ocean seeps and vents, microbial processes form the base of the food web and provide energy to support all of the complex metazoans, protozoans, and sponges.
Chemoautotrophic microorganisms can be free-living or symbiotic. The most obvious and generally abundant free-living chemoautotrophs at cold seeps are the giant sulfur-oxidizing bacteria. These bacteria, often members of the genus Beggiatoa, are large and have filaments that can sometimes exceed 100 micrometers (100 micrometers is about 0.0001 m or 0.0039 in) in diameter. These giant filaments are visible to the human eye (fig 1) and resemble strings of spaghetti. For reference, most bacteria are quite small — 1 or 2 micrometers in diameter — making them invisible to the human eye. At cold seeps, the Beggiatoa come in two main varieties, white and orange (fig 1). Both varieties oxidize hydrogen sulfide and they appear to differ in their sulfide tolerance (i.e., they have different tolerances for hydrogen sulfide concentration; the orange can tolerate higher concentrations) and in their ability to use carbon sources other than carbon dioxide (the orange variety can even use simple organic carbon substrates like acetate as its structural carbon source). Beggiatoa and other physiologically similar bacteria oxidize hydrogen sulfide to gain metabolic energy. But, where does this hydrogen sulfide come from?

Fig. 2. Oil-stained core samples from a Gulf of Mexico cold seep. Click image for larger view and image credit.
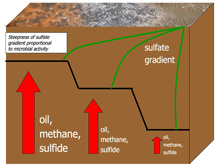
Fig. 3. Fluxes of oil and methane from deep sediments fuel consumption of sulfate and production of hydrogen sulfide. This hydrogen sulfide serves as a food source to chemoautotrophic bacteria like Beggiatoa (shown here on the sediment surface). Click image for larger view and image credit.
The BIOgeochemical Engine
At hydrothermal vents, hydrogen sulfide is present in the hot fluids venting from white and black smokers. This hydrogen sulfide is produced mainly by mantle-heated seawater interacting with ocean crust, a geochemical interaction. It is quite different at cold seeps: here, hydrogen sulfide is produced through microbial processes, by bacteria living in the shallow sulfate-rich sediments. These bacteria are sulfate reducers, which means they chemically convert or "reduce" sulfate to sulfide, using methane and oil oxidation to fuel the reaction. In the Gulf of Mexico, methane and oil seep together from the sea floor. Cores of mud from this region are often saturated with gas and stained by oil (fig 2). As oil and gas seep through the sediments, microorganisms living in the sulfate-rich zone convert the oil and methane to carbon dioxide, and in the process, they also convert sulfate, an abundant ion in seawater, to hydrogen sulfide (fig 3). The concentration of hydrogen sulfide builds up in the sediment and it flows upward towards the sediment surface. There, the hydrogen sulfide serves as energy (food) for Beggiatoa and other sulfide oxidizing bacteria. In the seep environment, it is microbial processes that provide the basic energy driving the deep-sea chemosynthetic ecosystems — a "BIOgeochemical engine."
On this expedition, we will examine the distribution of microorganisms, determine the patterns of microbial activity, and evaluate microbial diversity using sophisticated molecular biological techniques. Together, these data will help us to better understand how microbes and microbial processes form the energetic foundation of deep-sea chemosynthetic ecosystems.
References Cited:
Whitman, W. B., D. C. Coleman, and W. J. Wiebe, 1998. Prokaryotes: The unseen majority. Proceedings of the National Academy of Sciences, U.S.A. 95: 6578-6583.